[ Lay Language Paper Index | Press Room ]
Catherine Carr, cecarr@umd.edu;
Yezhong Tang, ytang@umd.edu;
Jakob Christensen-Dalsgaard, jcd@biology.sdu.dk
Department of Biology.
University of Maryland,
College Park, MD 20742-4415
Popular Version of Paper 2pAB1, ”Evolution of sound localization in lizards”
Presented 1:05 p.m., Wednesday afternoon, November 28, 2007,
154th ASA Meeting,New Orleans, LA
It is a recent, surprising finding that the eardrum was invented independently in at least five of the major groups of land vertebrates – the frogs, turtles, lizards, birds and crocodiles, and mammals. The ancestor of today’s land vertebrates did not have a tympanic ear. The earliest four-legged vertebrates probably were totally aquatic, but when they moved onto land, they must have had an ear more like that of fish, without a functioning middle ear and eardrum and responding to sound-induced vibrations of the skull (a kind of bone conduction). Such an ear would already have had crude low-frequency sensitivity, but the emergence of an eardrum and middle ear would have increased the range of frequencies heard and the sensitivity of hearing. Surprisingly, all the major groups of land vertebrates seem to have developed eardrums independently during the Triassic, roughly coinciding with the emergence of sound-producing insects, and more than 100 million years after the emergence of the first four-legged ancestors.
Additionally, the early middle ears were not enclosed in middle ear cavities, but were exposed in the mouth cavity. They were therefore acoustically coupled through the mouth cavity, making them inherently directional by a principle similar to the design of directional microphones. This is because such coupled ears work like pressure difference receivers – that is, the sound waves can reach both the outer and inner surface of the eardrum, amplifying or canceling its motion depending on the phase differences between the sound arriving at the surfaces of the eardrum. Recent vertebrates like frogs and lizards have retained part of this system. Most have wide Eustachian tubes, and sound waves may thus travel from one eardrum to the back surface of the other eardrum, creating an inherently directional pressure difference receiver. In lizards sound travels virtually unimpeded from one eardrum to the other, generating the largest directionality in any terrestrial vertebrate: up to a 50-fold directional difference as shown by laser vibrometry measurements of eardrum movement. Our recordings from auditory nerve fibers in lizards confirmed the prediction from the laser experiments and showed strongly directional responses. We were also able to show that the responses in the brain were directional. The function of the auditory portion of the lizard brain may be to sharpen the directional responses by binaural comparison, a relatively simple operation since the tympanic input is strongly lateralized. This probably provides enough information to orient the animal robustly towards a sound source.
Figure 1. Light shining through the Gecko head, from one ear to the other.
This is not how our ears work. In most modern mammals the middle ears are firmly enclosed in middle ear cavities, only communicating with the mouth cavity through Eustachian tubes that are so narrow that hardly any sound is transmitted from the mouth to the other eardrum. Our ears are therefore non-directional pressure receivers. We can compute directionality by comparing interaural differences due to head shadowing or the time of arrival of sounds at each ear. The closure of the middle ear cavity in the mammals and some birds seems to have occurred later in evolution, and may have profoundly changed the operation of the ancestral ear by decoupling the eardrums and leading to a requirement for additional neural computation of directionality in the brain. Most likely the closure of the middle ear cavities were caused by selection pressures to protect the middle ear, either mechanically or from respiratory noise.
|
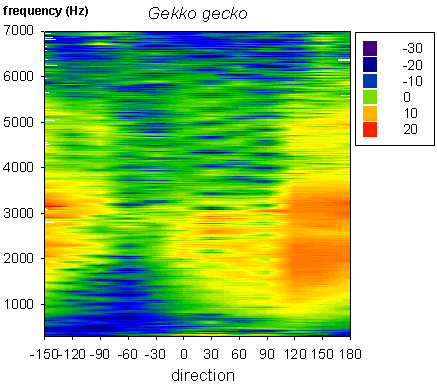
Fig. 2 Laser vibrometry measurement of directional sensitivity in the gecko. Positive angles are on the side of the measured eardrum, 0 is the frontal direction. Note the strong directional difference from 1 to 2 kHz |
|
Fig. 3. A cast of the mouth cavity of a gecko. Note the large, open middle ear cavities (arrows). |
|
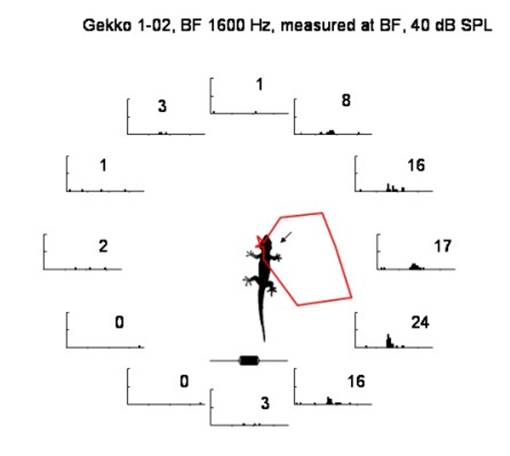
Fig. 4. Directional sensitivity in the gecko auditory nerve. Activity is recorded in a single nerve fiber in the right auditory nerve in response to directional (free-field) sound. The figures are histograms showing activity elicited by a 1.6 kHz tone burst. Number of action potentials are shown for each direction and plotted as a polar plot (red curve). |