
Lee A. Miller lee@biology.sdu.dk
Institute of Biology
University of Southern Denmark
Campusvej 55
5230 Odense M
Denmark

Popular version of paper number 2pAB1
Presented at 1:30 p.m. on Tuesday, November 11, 2008
156th Meeting of the Acoustical Society of America, Miami, FL

![]()
Picture 1-harbor porpoises Freja and her newborn, Frigg. Picture from August 2007, Fjord@Bælt, Kerteminde, Denmark
Echolocation, or the ability to orient using echoes from emitted sounds, was first discovered for bats in 1938 by Pierce and Griffin (1). During the 40’s and 50’s Griffin (2) described, among many other things, how free flying bats detected and captured their insect prey with the help of echolocation. During the same time others discovered insects that could hear and evade bats. It took more time to discover the only other group of animals using echolocation, the toothed whales (3). This was partly due to the difficulty of working in an aquatic environment and the availability of study animals. The first description of how dolphins in captivity captured fish appeared in 1972 (4). However, we still do not know much about how free ranging toothed whales find and capture prey using echolocation
Studying prey capture by toothed whales in captivity can give us insight into how they use echolocation for this task, and also shed light on how this might be accomplished by free ranging animals. But, which animal to chose? As the Danish Nobel Laureate August Krogh once wrote; for every biological question there is an optimal organism. We think the harbor porpoise, Phocoena phocoena, satisfies the August Krogh requirement. It is small and easily handled, numerous (at least in Danish waters), it can be kept safely and healthy in relatively small pools, they can be fairly easily trained, and they even breed in captivity (see picture 1).
Two harbor porpoises were trained to capture live (and dead) fish at our facility, Fjord&Bælt, in Kerteminde, Denmark. One actually did this when blindfolded. Their echolocation clicks are out of our hearing range, being at about 130 to 150 kHz. They are also very brief; about 100 µs long (see pictures 2 and 3).

Picture 2-A harbor porpoise echolocation click shown in time and relative sound amplitude.
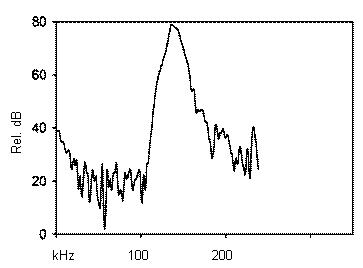
Picture 3-Frequency spectrum of a harbor porpoise click. Maximum sound amplitude at ~130 kHz.
They emit their echolocation clicks continuously, day and night, for orientation and searching for prey. When they find prey the pattern of clicks changes from slower searching rates to higher rates of clicks (shorter intervals) when approaching the fish (picture 4). The click interval is kept fairly constant at about 55 ms (red line in picture 4) just before the interval continuously shortens (rate increases). The highest rates (>600 clicks/s) occur just before capture. You can hear a facsimile of picture 4 in audio 1 (reduced by 22.7 times so we can hear the echolocation signals).
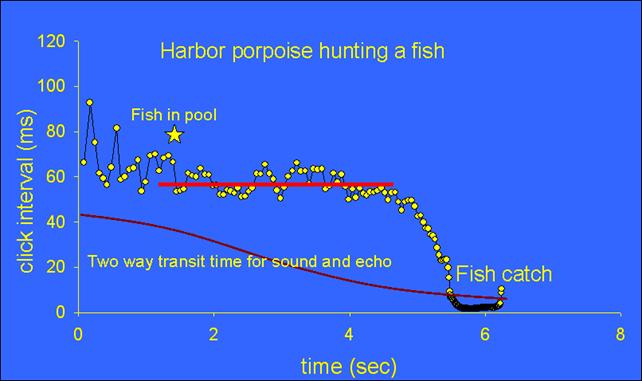
Picture 4-Harbor porpoise echolocation click intervals during hunting and catching a fish. (Verfuß et al. submitted)
Also, while the harbor porpoise approaches the target it reduces the loudness of its emitted echolocation click by about half for each halving of the distance to the target (see picture 5). This means that the sound level hitting the target remains nearly constant as the echo level at the porpoise’s ear increases.

Picture 5- Freja reduces the amplitude of her echolocation signal by about half (6 dB) for each halving of the distance to the target.

WATCH: By clicking on the picture above, you can see and hear how a harbor porpoise catches a fish. The sound comes via a detector so we can hear it, and the speed is slowed down to 1/10, since the capture goes very fast. Hear how the rate increases as the porpoise closes in on the prey.
What can we conclude from these studies? First, the pattern of echolocation signals of insectivorous bats and our harbor porpoise are similar although bat signals are longer in time. For comparison here is a recording of a bat (the New Zealand long tailed bat, Chalinolobus tuberculatus, reduced by a factor of 32) catching an insect in mid air.
This indicates that there are some basic rules for echolocation shared by these two groups of mammals living in vastly different environments. Secondly, we know of some fish in the herring family that can detect and respond to the echolocation signals of toothed whales. However, we do not know if interactions exist in nature between fish that can hear echolocating tooth whales and the echolocater (5), like those documented between bats and tympanate insects (6). But, if so the harbor porpoise may have an advantage by keeping the interval between signals nearly constant after detecting the fish while keeping a nearly constant incident sound level on the fish. This could conceal the echolocating porpoise from detection by the fish. This is, of course, just a postulate, but the latter seems true for bats too. Thirdly, results from captive porpoises will help interpretation of recordings from toothed whales tagged with various types of acoustic tags (7, 8).
There are naturally many questions still to be answered using animals in captivity. For example how robust is the harbor porpoise echolocation system to interference? How do they deal with unwanted echoes (clutter) and noise in the environment? Can they identify prey based on echolocation alone? How good are they at detecting fishing nets, a major cause of incidental death of harbor porpoises?
Acknowledgements: For their comments, discussions and support I would like to thank: Ursula Verfuß, German Oceanographic Museum, Stralsund, Germany, Magnus Wahlberg, Fjord&Bælt, Kerteminde, Denmark, Kristian Beedholm, Aarhus University, Århus, Denmark, Jonas Teilmann, National Environmental Research Institute, Roskilde, Denmark, Tomonari Akamatsu, National Research Institute of Fisheries Engineering, Japan and all the students who contributed to numerous projects. I thank the ASA for organizing the special session in honor of Sam Ridgway and for their invitation to attend. The funding agencies are the Danish National Research Foundation and the Danish Research Council for Natural Sciences.
References:
1) Pierce GW, Griffin DR. 1938. Experimental determination of supersonic notes emitted by bats. Journal of Mammalogy19, 454-455.
2) Griffin DR. 1958 Listening in the Dark. New Haven: Yale University Press.
3) Schevill WE, Lawrence BD. 1956. Food finding by a captive porpoise (Tursiops truncatus). Breviora, Mus.Comp.Zool, Harvard University53, 1-15.
4) Morozov VP, Akopian AI, Burdin VI, Zaitseva KA, Sokovykh YA. 1972. Tracking frequency of the location signals of dolphins as a function of distance to the target. Biofisika17, 139-145.
5) Mann DA, Lu Z, Popper AN. 1997. Ultrasound detection by a teleost fish. Nature389, 341.
6) Miller LA, Surlykke A. 2001. How some insects detect and avoid being eaten by bats: The tactics and counter tactics of prey and predator. BioScience51, 570-581.
7) Madsen PT, Johnson M, Aguilar de Soto N, Zimmer WMX, Tyack PL. 2005. Biosonar performance of foraging beaked whales (Mesoplodon densirostris). Journal of Experimental Biology208, 181-194.
8) Akamatsu T, Wang D, Wang K, Naito Y. 2005. Biosonar behaviour of free-ranging porpoises. Procedings of the Royal Society of London B272, 797-801.